生物化學與分子生物學/腦代謝的某些特點
| 醫學電子書 >> 《生物化學與分子生物學》 >> 神經組織生化 >> 腦代謝的某些特點 |
| 生物化學與分子生物學 |
|
|
一、能量供應(Energy Supply)
腦的活動瞬息萬變,需要大量能量的及時供應。腦細胞本身的生物高分子(核酸及蛋白質)的合成以及神經遞質的合成與釋放固然都是耗能的過程,但這些尚不足以說明為什么腦細胞的功能活動較之其他組織細胞要消耗更多的能量。腦的能量消耗主要在于經常不斷地把Na+泵出細胞外,使去極化(depolarization)后的膜迅速恢復膜電位,以維持神經的興奮和傳導。腦的代謝率(metabolic rate)是很高的,它可以用單位時間的耗氧量(oxygenconsumption)和基質消耗量或產物生成量作指標來表示,如表14-1。
表14-1 正常青年男子的腦血流量和代謝率
| 速率 | ||
| 每百克腦組織 | 全腦 | |
| 血流量(ml/分) | 57 | 798 |
| 耗氧量(ml/分) | 3.5 | 49 |
| 葡萄糖消耗(mg/分) | 5.5 | 77 |
| CO2生成(ml/分) | 3.5 | 49 |
腦血流量占心輸出量的15%,耗氧量占全身總耗氧量(約250ml/分)的20%,然而腦的重量只不過占體重的2%。分析流入和流出腦組織血液的化學成分(動靜脈差法)發現,除了葡萄糖外,其他可作為能源的物質沒有明顯的減少。由上表可見每百克腦組織每分鐘產生的二氧化碳和消耗的氧均為3.5ml(156微克分子),呼吸商(respiratoryquotient)為1。再者,按化學計算(1克分子葡萄糖完全氧化要消耗6克分子的氧),葡萄糖的消耗率還稍高于氧的消耗率。
這些事實綜合起來說明,腦組織是以葡萄糖的氧化來供能的,甚至可以說,至少在正常條件下,腦組織唯一利用糖作為能源。因為腦中糖原含量很少(小于0.1%),所以必須依賴血糖的供應。雖然腦組織還可以利用酮體,但必須以低血糖為前提,例如在饑餓引起酮血癥(ketonemia)的情況下。如果血糖和血酮體均增高時(糖尿病酮血癥),腦仍然優先利用葡萄糖以供能。有人認為,腦利用酮體作能源是對饑餓時低血糖的適應,長期慢性饑餓的病人,腦的耗氧量的一半可用來氧化酮體。
腦細胞含有完整的糖酵解(EMP)酶系,已糖激酶活性約為其他組織的20倍。但是即使最大程度地發揮糖酵解的作用也不能滿足供能的需要,而必須依賴糖的有氧氧化。所以氧的供給一刻也不能中斷。由于腦組織主要依賴糖的有氧氧化供給能量,所以它對缺糖和缺氧均極敏感。血糖下降50%即可致昏迷,而中斷(流向腦的)血流幾分鐘就可引起死亡。臨床上使用大劑量胰島素做為治療手段也可以引起昏迷。胰島素對腦的這種影響,現在認為是間接作用的結果,也就是因為胰島素降低了血糖之故。因為胰島素是大分子物質,它不能透過血腦屏障,這和對其他組織或周圍神經組織的作用不同。在那些組織,胰島素的作用可能主要是直接影響其對葡萄糖的攝取。
腦內ATP的水平甚高,它的合成和利用均很迅速。據測定,腦內ATP末端磷酸基的半數更新時間平均只有約3秒鐘,腦組織的磷酸肌酸(CP)水平比ATP還要高,它可看作是ATP末端高能磷酸鍵的一種貯存形式。在磷酸肌酸激酶(CPK)的催化下,ATP和CP可相互轉變。
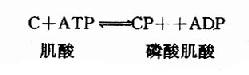
這是一個等能反應,因此在相互轉變中不致于有能量的丟失。腦中CPK活性較強,有利于能量的貯存和動用。此外,腦組織中還有活性較強的肌激酶(myokinase),它可催化下列反應。此反應的意義在于“挖掘潛力”,即充分利用ATP中的兩個高能磷酸鍵。
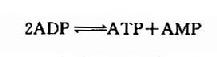
上述兩種激酶都是維持腦內高水平ATP的有力保證。
二、類脂的組成和代謝
除脂肪組織外,腦是全身含脂類最多的組織,但脂肪組織主要含甘油三酯(貯存脂),而腦組織中的脂類幾乎全是類脂。腦干重的1/2是脂類,這是就全腦平均而言,如果分別測定腦灰質和腦白質的化學成分,就會發現灰質含水份和蛋白質較多,脂類僅占干重的1/3;而白質中的脂類含量較多,約占干重的55%。
表14-2 正常成人腦的脂類組成
| 成分 | 灰質 | 白質 | ||||
| 溫重(%) | 干重(%) | 總脂(%) | 溫重(%) | 干重(%) | 總脂(%) | |
| 水 | 81.9 | 71.0 | ||||
| 總脂 | 5.9 | 32.7 | 100 | 15.6 | 54.9 | 100 |
| 總磷脂 | 4.1 | 22.7 | 69.5 | 7.2 | 25.2 | 45.9 |
| 膽固醇 | 1.3 | 7.2 | 22.0 | 4.3 | 15.1 | 27.5 |
| 總半乳糖苷脂 | 0.4 | 2.4 | 7.3 | 4.1 | 14.5 | 26.4 |
| 總神經節苷能 | 0.3 | 1.7 | 0.05 | 0.18 | ||
由上表可見,以濕重計,腦白質中脂類含量約三倍于灰質。這種差別主要是由于白質中的神經纖維外被以髓鞘(myelin sheath),而髓鞘的脂類可高達干重的70-80%。
腦中的類脂主要用以構成神經元(neurone)的質膜和髓鞘。這些膜性結構與其他組織細胞的膜結構有共同之處,即都是由類脂與蛋白質構成的復合物,但在類脂的組成和代謝上亦有一些特點。尤其是髓鞘,它含有某些特殊的類脂成分,這些成分或者僅見于髓鞘,或者髓鞘中含量較多,而在其他組織中則較少見,例如縮醛磷脂(plasmalogen)和腦苷脂(cerebroside)。
腦不從血中攝取脂肪酸,本身也不含游離脂肪酸,那么組成這些類脂的脂肪酸是從哪里來的呢?實驗證明,腦中的脂肪酸和膽固醇都可由乙酰CoA合成,而乙酰CoA的主要來源還是葡萄糖。
髓鞘形成(Myelination)之前的未成熟的腦組織含膽固醇和磷脂較多,而含腦苷脂極少,腦苷脂合成酶系的活性也極低。當髓鞘形成時,此酶系的活性升高,腦苷脂的含量亦相應增多,髓鞘形成與神經系統的發育和功能密切相關,而髓鞘脫落(demyelination)是神經系統疾病的重要的病理改變之一。髓鞘的代謝特點是正在進行髓鞘形成時代謝很快,一旦形成之后就變得很慢,成為體內最穩定的一種結構。據認為,這是由于髓鞘缺乏催化類脂分解代謝的酶系。已經形成的髓鞘,除了個別成分(如三磷酸肌醇磷脂)有較高的更新率外,其他磷脂和膽固醇等的更新率均甚低。
三、谷氨酸的代謝與功能
腦的游離氨基酸組成與血漿有很明顯的差別,這是由于血腦屏障的特點和腦本身氨基酸代謝特點造成的。腦和血漿中某些游離氨基酸的含量對比如表14-3。
表14-3 人腦和血漿中某些游離氨基酸的含量
| 氨基酸 | 腦 | 血漿 |
| 微克分子/克 | 微克分子/毫升 | |
| 谷氨酸 | 10.6 | 0.05 |
| N-乙酰天門冬氨酸 | 5.7 | - |
| 谷氨酰胺 | 4.3 | 0.7 |
| γ-氨基丁酸 | 2.3 | - |
| 天門冬氨酸 | 2.2 | 0.01 |
| 色氨酸 | 0.05 | 0.05 |
由表14-3可見,腦中游離氨基酸以谷氨酸(Glu)含量最高,它比其在血漿中的濃度要高出200倍以上。谷氨酸、谷氨酰胺(Gln)和γ氨基丁酸(GABA)三者含量總和約占腦中游離氨基酸總量的一半。所以,在腦的氨基酸代謝中,谷氨酸占有重要位置。
然而,谷氨酸難以通過血腦屏障,腦內谷氨酸來源于自身的合成,同位素示蹤實驗表明腦內谷氨酸合成的原料是葡萄糖,它來自血糖。葡萄糖進入腦細胞后先轉變成α-酮戊二酸(α-KG),后者可在谷氨酸脫氫酶的催化下轉變成谷氨酸,亦可經轉氨基作用生成谷氨酸,一般認為后一途徑更切合實際,因為谷氨酸脫氫酶(GDH)催化逆反應時KmNH4+為8mM,遠高于細胞內氨之濃度。谷氨酸在谷氨酰胺合成酶的作用下與氨結合成為谷氨酰胺,這是一個耗能反應(消耗ATP),腦中谷氨酰胺合成酶的活性強,其KmNH4+僅為0.39mM。所生成的谷氨酰胺,與谷氨酸不同,可以通過血腦屏障而進入血中,這樣,腦組織從血中攝入葡萄糖,通過代謝,還血液以谷氨酰胺,清除了腦中的氨,以免氨的積存危害腦的功能。
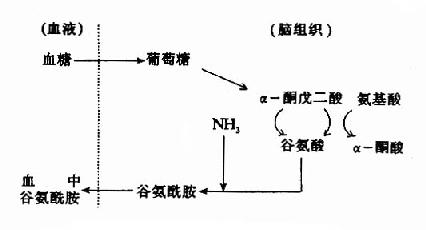
圖14-1 腦中谷氨酸代謝與氨的消除
腦中谷氨酸代謝的另一個特點是脫羧生成γ-氨基丁酸(又名γ-氨酪酸GABA),催化此反應的酶是谷氨酸脫羧酶(GAD),它需要磷酸吡哆醛作輔酶。GABA是一種抑制性的神經遞質,僅見于中樞神經系統。腦內GABA主要貯于灰質,特別是紋狀體、黑質、小腦的齒狀核等處。
GABA對中樞神經元有普遍性抑制作用。1963年曾有人提出,GABA能作用于突觸前神經末梢,減少興奮性遞質的釋放,從而引起抑制。這種效應稱為突觸前抑制(presynaptic inhibition)。GABA在脊髓中的作用就是以突觸前抑制為主。在腦內則GABA主要是引起突觸后抑制(postsynaptic inhibition)。睡眠時皮層釋放GABA增多,因此有人認為GABA可能與睡眠、覺醒的生理機能有關。
|
|
|
| 圖14-2a 腦中TCA循環和GABA代謝旁路 | 圖14-2b GAD和GABA-T的作用 |
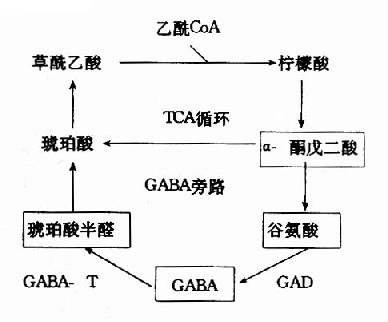
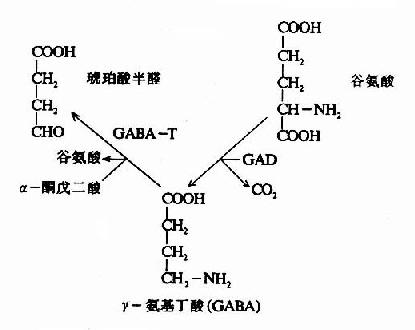
在神經元胞體和突觸(synapse)的線粒體內含有大量的γ-氨基丁酸轉氨酶(GABA-T),它可催化GABA與α酮戊二酸之間的轉氨作用,生成琥珀酸半醛(succinic acid semialdehyde)和谷氨酸。這可看作是GABA滅活的一種方式。GABA-T也是需要磷酸吡哆醛作輔酶,但與GAD比較,它同磷酸吡哆醛的親和力大,所以當體內維生素B6缺乏時,主要影響GAD的活性。例如,使用異煙肼治療結核病時,由于異煙肼能與維生素B6(吡哆醛)結合成異煙腙(isoniazone),加速維生素B6從尿中排泄,引起腦組織內維生素B6濃度下降,GAD活性亦下降,結果GABA的合成受阻,容易使中樞過度興奮而發生抽搐等癥狀。所以長期使用異煙肼時應合并使用維生素B6。此外,臨床上對于驚厥、妊娠嘔吐的病人,也常使用維生素B6,其道理也是提高腦組織內GAD的活性,使GABA生成增多,中樞抑制相對加強。
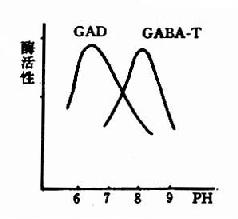
圖14-3 GAD與GABA-T的作用最適pH
GABA經轉氨作用后的產物琥珀酸半醛可脫氫生成琥珀酸,后者進入三羧酸循環而被氧化利用。因此,與腦組織中的三羧酸循環相連系,存在著一條GABA代謝旁路(GABa shunt)。
谷氨酸脫羧酶與γ-氨基丁酸轉氨酶的協同作用對保持腦中GABA一定濃度有重要意義。兩種酶的最適pH不同,GAD的最適pH為6.5,而GABA-T則為pH8.2。(圖14-3)由此可見,腦細胞內pH稍有變動就可明顯改變這兩種酶的活性對比。當酸中毒時,腦中GAD活性增強而GABA-T活性減弱,可致腦中GABA水平上升,呈現中樞抑制;反之,當堿中毒時腦中GABA-T活性增強而GAD活性減弱,腦中GABA水平下降,易于發生痙攣。
尚須指出,谷氨酸對神經中樞有興奮作用,而其脫羧產物GABA卻有抑制作用,所以谷氨酸的代謝與中樞的興奮和抑制調節有關。此外,通過GABA代謝旁路,也把腦的氧化代謝與興奮抑制功能聯系起來了。
| 關于“生物化學與分子生物學/腦代謝的某些特點”的留言: | |
|
目前暫無留言 | |
| 添加留言 | |